BIOGEOGRAPHY/PALEOECOLOGY,
Fall 2018
SECOND PROBLEM SET
PHYLOGENIES:
1. Here are 14 species of a type of organism known as caminalcules..
Your job is to propose a phylogeny linking them. Assume the
group, as a whole, is 'good' monophyletic group, and all of the types
shown represent distinct, living populations. Proceed by choosing
potentially useful 'characters' and identifying two or more
'character-states' (for example, number of eyes might be a character,
and character-states could be 0, 1, or 2. OR you might say
character is just 'eyes' and states are 'present' or 'absent'; then
number of eyes could be used as another character only for those with
eyes present.
- Choose at least
six characters to work with; list them with the observed states.
- Decide how you will interpret states as plesiomorphic
or apomorphic (explain your reasoning) (ASSUME that these
are vertebrates remotely related to amphibians -- that is, you
could use
a salamander or a frog as an 'out-group' to help identify
plesiomorphic states).
- Use shared apomorphies ('synapomorphies') to construct an 'optimal'
cladogram/phylogeny, trying to minimize the number of state conversions
(evolutionary events) required. (i.e., a mazimum
parsimony tree). Identify end-points ('present time') on your
cladogram by the
numbers below.
- Identify, on your cladogram, branches where state
changes are inferred.
- Suggest a division of these 14 species into taxonomic
sub-groups (you can give them names if you want).
- If your phylogeny involves convergences or
reversions, point these out.
- Thinking of your cladogram as a
phylogenetic hypothesis, discuss
what kinds of additional information (beyond the observed morphology of
the creatures) could allow you to test it more rigorously. (Keep
in mind that there might be other topologies that are as
parsimonious -- or nearly so -- as yours, and parsimony is a somewhat
arbitrary criterion. Also, that different assumptions about
what's ancestral/derived might change everything...)
(you can hand in your cladogram on paper if you prefer.)
Lots of ways to answer this, of course, but here are some important points...
-
assignment of plesiomorphic(ancestral) vs. apomorphic (derived) 'state'
for particular characters: this is most powerfully done using
'out-groups'; trait-states shared with out-group organisms are usually
assumed to be plesiomorphic for the clade under consideration.
SO, if
the out-group is 'frogs', then you'd assume having a head, 2 eyes, 4
legs with toes are ancestral/plesio conditiosn. Thus, eyeless
species,
here, would represent a derived condition... It is NOT a very
'safe'
approach to assume the 'simpler' = 'ancestral' (e.g., eyeless is
ancestral, eyes = derived); selection causes loss of structures/traits
as often as it adds them (probably more often). How common the
trait
is across the group is also not a good indicator of ancestral vs.
derived (think about it). That means that it's very hard to
assign
some traits (like body shape or marking patterns here - both of unclear
relationship to outgroup AND things that change rather easily in
evolution); often that's done by working with
the 'known' apomorphic states and trying to organize tree so that there
are as few as possible transitions of other traits, THEN deciding which
state is ancestral.
- Note that question specifies that all of these things are living species, so your tree should NOT assign one as an ancestor of another; they should all be positioned at 'twig tips' in the tree...
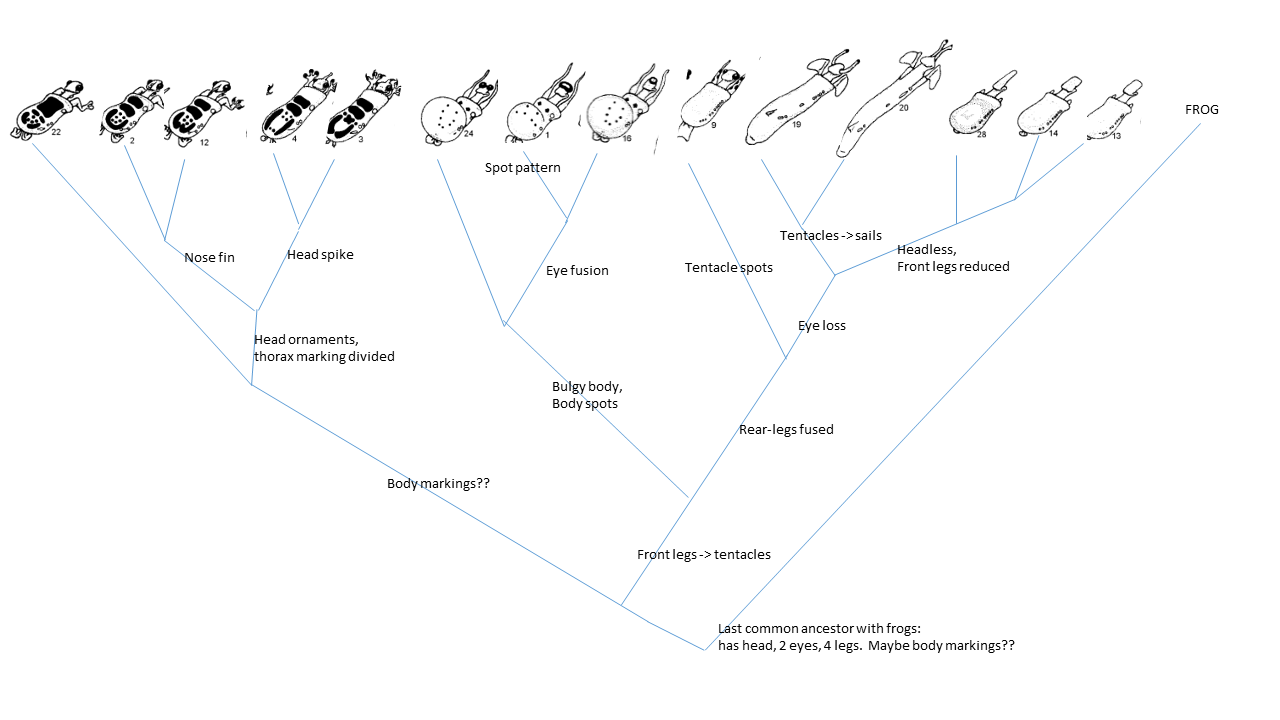
-
Here's one example of a possible cladogram; depending on some judgments
about sequence of trait loss or acquisition, others are possible. Note
that this one includes: convergence on clawed toes (12 and 2) OR
secondary loss of claws in 4; ambiguous arrangement of 13,14,28...
-
I might assign to three genera, left to right: tetrapods
with body
ornaments; tentacled shoe-bodies with spots; and legless wonders.
(Number 9 becomes EITHER a separate genus or goes with the legless ones
to maintain monophyly)
NOTE that 22 could be set up as a 'basal group' , sibling to all
other
caminalcules and ancient common ancestry, so would have to be yet
another genus (this would
be parallel to the two 'singleton' branches in squirrel tree in next
question).
 .
.


2.
The diagram below is a proposed 'family tree' or cladogram for
squirrels. (J.M. Mercer and V. L. Roth,. 2003. The Effects of Cenozoic
Global Change on Squirrel Phylogeny. Science 299:1568-1572) Each
terminal branch is one living taxon of squirrels (except for
Aplodontia, which is weird rodent called a mountain beaver, and is used
here as an 'out-group'). Remember that no extinct taxa are shown.
Don't worry about the technological details in the caption; just assume
that the relationships shown are correct. Answer two of the
following:
A. The tree shows two 'basal' lineages (or
branches) – Sciurellus and Ratula – that originate very near the 'root'
of the squirrel tree – their common ancestry with other squirrels is
quite ancient ; each of these two genera has only one species. the
three more recently diverging branches each have many genera. The
researchers suggest that these two ancient 'monotypic' lineages
required an explanation because it seemed odd that a single species
would persist so long 1) without going extinct and 2) without
diversifying like the other branches. Offer an explanatory
hypothesis or two.
B. The researchers say the big
diversification of squirrels was rather sudden and more or less
simultaneous in all the major branches, and happened at the end of the
Eocene at a time of "significant climate change and extinction.”
Why would it make sense that such a dramatic (apparent) diversification
within a lineage should coincide with such events?
C. The little pictures indicate branches that are tree-squirrels,
ground squirrels (like chipmunks; that's the little prairie-dog-like
symbol in group IV), and flying squirrels; ground and flying squirrels
each occur in only one main branch; tree squirrels occur in all
three. What do you think that tells you about the history of
these three styles of being a squirrel?

A.
At least four general possibilities; H1: what these two critters
(genera actually; look them up; they're cool) do is very
successful but very ecologically specialized; there's only one way of
doing it, so not a lot of divergence. This might well combine
with H2: they've been geogrpahically
'stranded' in some isolated situation that prevented
sufficient genetic isolation for speciation within the lineage. (NOTE
that this does not imply they've stayed the same all that time; they
could have been changing a lot, just not splitting...) H3: An
almost
opposite line of reasoning; they are super-generalists that can live
almost everywhere squirrel-able, so no opportunity for
isolation/speciation -- like, say, mountain lions (but this isn't
consistent with actual range). Or, H4: the lineages
WERE more diverse, but other species/genera belonging to their
sub-clades have
gone extinct; Ratula and Sciurellus, then, would be remnants of
previously more diverse groups (like Homo sapiens).
NOTE that these hypotheses don't necessarily imply that the
modern species are particularly ancient (except maybe H3). .
B. Several possibilities again; H1:
mass extinction opens up a lot of ways of making a living through
variations on surviving lineages, previously outcompeted by other
organisms; these can now get a foothold and gradually become more and more
distinct and specialized = new adaptive radiation [this
REQUIRES that there's an opportunity for populations to diverge
adaptively with subsequent development of RIMs;,
OR H2: big changes
in environment can, even without extinction of existing biota,
create new opportunities that would
foster adaptive radiation (maybe forests became dominant where they
were previously rare, and being a squirrel suddenly makes a lot of
sense...), OR H3: What changes is opportunity for isolation; the climate change caused fragmentation of squirrel
habitat (e.g., extensive forested landscape becomes fragmented -- this
is consistent, for example, with a drying climate), generating lots of
isolated populations that diverge in allopatry (adaptive/selective
processes not even necessarily required at first)..
C. DISREGARDING Ratula and Sciurellus, there are three ways of
interpreting this in terms of plesiomorphic and apomorphic traits: 1)
the common ancestor of all squirrels was a tree squirrel;
ground-sq and flying-sq life-histories originated one time each, on
separate 'branches' of the big clade (but --
a complication -- it looks like there's a 're-invention' of
tree-sq-ness on clade IV, so that would add a change for three total...);
2) overall ancestral trait is
'ground-sq' requiring origin of tree-sq-ness on each of the three main
branches independently and flying-sq-ness once; 3) flying is ancestral
state, requiring similar pattern to 2. Option 1 requires the
fewest changes from the common ancestor of all squiirels (even though
it may require one convergent reinvention of tree-dwelling -- it is the
most parsimonious story. It's FURTHER supported by the two 'basal
groups' -- Sciurellus and Ratula -- being tree squirrels (maybe
strongest point...).
OTHER STUFF (you can do THREE of these -- or do all four and I'll 'score' the best 3):
3. Isolated, small
populations with ranges restricted to relatively small, strictly
bounded habitat patches (like an island, or a desert oasis) appear to
undergo more rapid selective/evolutionary change than do large
species/populations distributed over large geographical areas. Offer at
least one hypothesis for why this should be so. (hint: you might
consider relationships between selective evolutionary change and
non-selective change like ‘gene flow’ or genetic drift -- genetic
change that's not directly related to adaptedness.)
This needs to be in terms of changes in genetic make-up of populations – that is the definition of evolutionary change. Several possible lines of thinking can contribute:
- smaller populations change more rapidly through
genetic 'drift' simply because they're smaller: a random mutation in
one individual (because that individual is, effectively, a larger
proportion of the population) is more likely to spread through the
population 'accidentally' (regardless of selection); similarly, a
particular allele is more likely to be lost from existing gene pool for
similar reasons. Note, however, that drift reduces
genetic diversity and so, as the population becomes more uniform,
potential for further evolutionary change through any mechanism is reduced.
- inbreeding is more likely in smaller
populations; this has the same consequences as genetic drift – more
rapid (and essentially random) loss of genetic diversity for a time (until no heterozygosity remains).
Both of these are 'non-selective' processes; they are not
driven by natural selection – they are not due to heritable traits
driving differences in reproductive success (that is, differences in
fitness). There are also arguments involving selection:
- If isolated populations are established by
colonists from elsewhere, island habitats may well be somewhat
different than habitats from which island is colonized; colonists would
be subject, then, to new (directional) selective
pressures, different traits would be favored than on the 'mainland',
and, because there's little or no gene-flow from 'main' population to
counteract this divergent selection, it could be pretty rapid.
(Same arguments if isolation is created by vicariance -
'splitting' of existing range --, and lack of founder effect might mean
more initial genetic variation for selection to act on = faster change
-- IF the isolated bit has different enough selective regime than main
range (which is likely, as such vicariance events tend to be at edges
of ranges. Think about that...)
- By definition there'd be less gene flow
between isolated and non-island populations (less movement of
individuals
between than within the two). Thus, whatever traits are 'selected
for' on the island would be less subject to 'dilution' by gene flow
(immigration) from the mainland, so effects of selection will be more
rapidly 'fixed' within the island population. Another way of saying
this: selection pressure is likely to be more uniform across a small,
isolated population where, even if a locale
within main range is environmentally distinct, has similar
selection pressures as the 'isolate', gene flow within the main range
will tend to reduce the possibility that a local population could
diverge evolutionarily (THIS IS AN IMPORTANT THING to think about).NOTE that this questions was not about speciation; more rapid evolutionary change MIGHT increase likelihood of speciation, but that does not necessarily follow.
4. Dawkins
makes a clear distinction between 1) the tendency for Darwinian natural
selection to produce specialized 'survival machines' that serve
as vehicles for the replicators (genes) that build them, and 2)
the potential role in the grand picture of a property he calls 'evolvability'
-- the potential, within a lineage or clade, for generating new traits
on which selection might act.
A)
If individual lineages within a larger taxon or clade tend to become
more adaptively specialized over time, what are the likely consequences
for particular species lineages over time? What effect, if any,
might this process or tendency have on the replacement of one major
group by another?
B) What properties might make an organism "good at evolving"? (these might be at genetic or phenotypic level...).
C)
Could the property of 'evolvability' (the potential for rapid
evolutionary change) confer fitness benefits on individual organisms?
If not, is there any reason to think that there might be a
tendency, over time, for life forms to become more 'evolvable'?
A)
There's probably a fundamental trade-off between specialization
and capacity to deal with changing circumstances (in both ecological
and economic systems...). More specialized lineages are likely
more vulnerable to extinction. The answer to second part would
depend on whether ALL species lines within a higher taxonomic group tended to become more specialized
; if they did, then big environmental changes might lead to loss of
such groups and their replacement by new 'radiations' from some more
generalist line. But, if the larger group (class, order,...)
retained some less-specialized lineages, there might continue to be
turnover within the group, but less chance of whole group being lost.
B)
The simplest: genetic variation within populations. There's no
evolving without that. So whatever would enhance genetic
variation within a population: higher mutation rate; high reproductive
rates (more offspring = more chanceof mutation); mating systems with
lots of out-breeding (or just sexual relative to asexual reprod);
polyploidy (more functional genes to mutate into something else, with
'backup').. Note that phenotypic
plasticity can have the opposite effect; if an organism can respond
developmentally/phenotypically to different environments without
changes in genotype, this can reduce potential for selection to act on
genome since 'adaptation to environment' is no longer due to genetic
differences as much...
C)
First part: possibly, but only under rather special circumstances.
The property that makes a lineage more 'evolvable' would have to
generally enhance probabilities of individuals leaving descendants OR of alleles 'causing' evolvability producing surviving copies of themselves.
Sex is good example; sexual reproduction reduces effective
fitness by half given constant number of offspring (any of your alleles
have only 50% chance of being in a specific offspring), so , for sex to
enhance individual fitness, the sexual individual in an asexual
population (never mind problem of finding a mate) would have to leave
twice as many offspring for same genetic fitness! This MIGHT be
the case if some aspect of the selective environment is changing in a
very rapid and unpredictable way... It's important
to differentiate between fitness of individuals within a population
from the likelihood of a species/line surviving.
5. Offer two general hypotheses for why there are no wheeled mammals
(except human extended phenotype); if flight had not already evolved in
mammals, would similar reasoning have applied for why there were no
flighted mammals?
A)
It''s not possible within the scope of mammalian anatomy/function (for
example, might be no way of arranging vascular or nerve tissues to pass
through a fully rotatory joint); B) it might be a design possibility,
but the selective pathway hasn't been available -- i.e., you'd have to
have a series of step-wise mutations/phenotypic changes EACH of
which enhances fitness.; C) perhaps it's both possible AND initial
steps have occurred, but they weren't selected for BECAUSE rolling
locomotion just wouldn't work well in mammalian habitats, and other
modes of movement would confer higher fitness. Neither of these
would be as convincing regarding potential for evolution of flight,
both because intermediate steps in converting leg to wing aren't as
challenging AND because we see flight have evolved independently in
other lines of quadruped vertebrates (at least two).
6. Nearly two million years ago, Homo erectus (or, according to more recent taxonomies, Homo of some pre-sapiens species
that should have a different name....) spread out of Africa across all
of southern Eurasia. By 100,000 years ago or so, Homo erectus was gone, and Homo sapiens occupied more or less the same region. Two general scenarios for this replacement have been proposed:
The 'Out-of-Africa hypothesis' proposes that H. sapiens originated in east Africa, where our species differentiated from ancestral H. erectus (or similar), and spread outward, replacing H. erectus populations (presumably through competition, but predation not excluded...) as they went. Thus, all H. sapiens share a common ancestor in the earliest H. sapiens populations in Africa.
The 'Multiregional hypothesis' suggests that H.erectus populations
throughout this range were gradually modified, by common selective
pressures and as a consequence of frequent immigration/gene flow among populations, so that H. sapiens developed in continuity with H. erectus throughout its range. In this scenario, the ancestors of, for example, east Asian H. sapiens would be largely or entirely east Asian H. erectus, and the most recent common ancestor of all H. sapiens would be much earlier, in the H. erectus lineage.
Address the following questions:
A)
Do either of these hypotheses seem more plausible in principle, given
what you understand of the processes of natural selection and
speciation (explain)?
B)
What sorts of archeological/paleontological evidence might be useful in
discriminating between the two hypotheses (i.e., do the hypotheses
yield different predictions in terms of expected patterns)?
C)
What sorts of genetic relationships among existing humans are
suggested, and how might these be used to assess the hypotheses?
A)
The MRH requires EITHER that selection would be similar enough from
East
Asia and Java to Africa (the range of H. erectus) that local selection
would produce sufficiently similar changes (and that there were
not significant mutations that arose only in one region) OR that there
was continuous and significant gene flow across that entire region (or
some combination of these). Not impossible -- but the OAH invokes
something more in keeping with conventional understanding of speciation
-- i.e., that local, allopatric populations are more likely to undergo
relatively independent evolution and those that discover 'new tricks'
are apt to then expand and displace their cousins. Additionally,
sustained genetic 'connectivity' over 2 million years might be not so
plausible. Most people
see the OAH as being more parsimonious -- calling for fewer 'special
circumstances' -- but this is always debatable.
B)
Most straightforwardly, OAH predicts fossils of intermediate forms ONLY
in Africa (which is the way most interpret the actual fossil record);
if intermediates turned up in East Asia, that would be a problem for
OAH. Same argument regarding tools/technologies. Conversely MH suggests that archeology should find
gradual transitions in anatomy AND in cultural development everywhere.
Cooccurence in place and time of distinct H. sapiens and H. erectus
fossils IN ASIA would be strong evidence against MH. Also, first
appearance of H.s. fossils should be progressively younger with greater
distance from africa under OAH: more simultaneous under MH..
C)
MH says our common genetic ancestry (MRCA) should be much older than
predicted by OAH. This is hard to test -- but MH would also suggest many more unique genetic
traits in non-African locations and similar levels of genetic diversity
throughout the range occupied by H. erectus; that is not the case. OAH predicts that
'deeper' genetic divergences would all have happened in Africa, so
higher diversity of genotypes likely in Africa, with older (as dated by
molecular clock) divergences than genotypes present ONLY in Asia.
(Note that this is true even if the clock is not well-calibrated
-- it's a comparativewithin our lineage...)