
FORESTS, FALL 2019
Question
set THREE
DUE
2.
Increased
movement of people and materials around the world has, in recent
decades, led to the introduction and establishment of many
non-indigenous species in our area (for example, about 1/3 of the
approximately 2100 plant species growing 'wild' in Vermont are the
result
of human introduction). Introduced species include plants,
animals, fungi, and microorganisms (including some that are pests or
pathogens). Many conservationists view such introduced organisms
as potential environmental threats, but views on this issue
differ.
A. Using
what you have learned about species interactions (competition and niche
theory),
suggest two or three ways in which introduced PLANTS might affect
regional natural communities and their diversity. Under what what
circumstances
would you regard introduction of a non-indigenous species as an
environmental 'threat' ? Explain your
reasoning (you may need to introduce some subjective valuations here;
that's okay). (You
can think in terms of any taxonomic group or guild of organism --
plants, animals, parasites, predators, etc. -- for purposes of
argument/illustration; if your arguments might apply more to some
groups than others, say so...)
B. All of the species that live in our area have dispersed here from
further south over the last ~14,000 years; before that this area was
ice-covered. Consequently,
ecological communities have gone through long series of transformations
as species expanded ranges following the retreat of the ice (and,
later, may well have 'contracted' their range northward). How
are these changes similar to or different from the consequences
of introductions of species from other parts of the world by human
agency?
This could be approached in several ways. For Part A: there are several possible avenues by which introduced plants could affect local communities and their diversity; if they use resources that aren't currently limited/limiting (i.e., occupy an 'empty niche' ) they might simply persist with little consequence except increasing diversity by one species. If they compete strongly with native species, they'll reduce abundance of one or more native populations and increase likelihood of extinctions, thus reducing diversity. They might change environment in other ways (e.g., changing soil properties or casting heavier shade) which could change which other species can coexist with them (potentially reducing diversity). The question about 'environmental threat' is, perhaps, trickier; the main issue here is to distinguish an 'effect' (like those mentioned alread) from a 'threat'. Is loss of native species or reduction in diversity good, bad, or neither? Unless you think ecological communities have their own goals and ambitions and purposes, that is up to you to determine -- it's a human evaluation, and that is important to recognize. Worth noting that some introduced species actually increase NPP/biomass or diversity; are these 'good' introductions?
For
B part, again, several possible lines of thought are potentially
relevant. One difference may be in the rate of new introductions
over time; at least recently numbers of human introductions has been
very high, and probably much higher than rates of new species arriving
by 'self-introduction' (although this may not be true for
birds...). Maybe more important as a 'meaningful' difference,
'natural' introductions through range expansions are likely to involve
shorter distances (and/or movements that don't cross 'barriers'), which
might mean that the interacting species are more likely to have had a
'coevolutionary history' of some sort.
3. This figure shows the
distribution of two species of cat-tails – Typha latifolia
and Typha angustifolia – over a range of depths of
water. Negative depth means out of (above) the water. (Cat-tails are the
dominant plant in the wetlands around the Dickinson Pond; both of
these species occur on campus). The upper graph shows situations
where both species occur together (in sympatry); the lower graph
shows distributions in situations where only one of the two species
occurs (allopatric). The vertical axis is a measure of abundance
(don’t worry about different values between the graphs; it's the relative abundance
of the two species that's of interest here).
Interpret the patterns observed in terms of fundamental and
realized niches for the two species, indicating the implied
competitive relationships. If the observed differences between the
two graphs are a result of interspecific competition, you might
hypothesize that competition is for either light or mineral
nutrients (since these are perennial wetlands, it’s
presumably not about water!). Cat-tails are rooted in the sediments,
and presumably obtain mineral nutrients through their roots. Briefly,
lay out an experiment to attempt to test these hypotheses.
Explain your methods, and what you would expect if the relevant
hypothesis is correct.
This is essentially the same scenario as the trout question on previous set, but with plants; range of depths inhabited in allopatry (bottom) would define fundamental niche (at least on the niche space axis defined by water depth). Fundamental niches are largely overlapping, but it looks like T. angustifoliahas a somewhat broader distribution in allopatry, or fundamental niche (could say it's more of a habitat generalist). (NOTE: some of you focused on depths of maximum abundance, which is okay, but the niche is really about the RANGE of conditions/habitat inhabited) In sympatry, there appears to be an asymmetrical competitive partitioning of overlap area in niche space: the generalist T. angustifolia seems to be inferior competitor over much of the area of overlap of fundamental niches (i.e., is excluded from deeper water in sympatry), but T. latifolia doesn't show much change in it's distributional range in sympatry; realized niche not much change for T.l., but substantially reduced for T.a. (which is now confined to deeper water). This is a common pattern; more 'specialized' species are typically better competitors in their range of tolerance than are more generalist species.. The usual assumptions apply; we don't know if a) the wetlands involved in the two graphs are otherwise generally similar, or b) whether other plant species (and competitors) might be influencing distributions differently in the two situations. To test a hypothesis concerning competition between two species you always have two approaches; you can remove/constrain one species and see if the other expands in number/distribution, OR you can change the availability of the putatively limiting resource and see if there's at least some (perhaps temporary) increase in the species thought to be limited by competition (although both would likely grow until the resource becomes limiting again OR some other resource becomes limiting). Here, you'd need to do experiments that would influence availability of light OR mineral nutrients separately from the other (fertlizing might be easy; changing how much light's available to the putatively inferior competitor would be tricky, but you could do it...)
II. ANSWER THESE THREE QUESTIONS ABOUT ECOSYSTEM PROCESSESProbably easiest to start with the suggested parallel with the Lake Washington story (I give hints for a reason!). Where an ecosystem 'switches' from one state with one dominant class of organisms to another state with a different dominant type of organisms, it is likely a consequence of changes in relative abundance of resource inputs and a change in limiting resource (this term is key; use it!) driven by competition. There's a shift in resource types involved here, too -- from 'high-carbon' and low-other-nutrient materials (carbohydrates) to materials with higher relative proportions of nitrogen (proteins). If fungi could operate at very low nitrogen, while bacterial decomposers can't, and if bacteria are better competitors for resources when they're not nitrogen limited, then the differences in compost-ecosystem chemistry described might drive a 'state change' much like what happened in the Lake (though the nutrients involved are C(energy):N instead of P:N). (ALSO, note that limiting resource arguments are always about relative amounts - proportions -- not just total amounts...). In brief; more wood chips, etc. in proportion to N-rich materials makes nitrogen limiting, favoring fungi.
The 'richer' (in nitrogen) compost environment that permits bacteria to outcompete fungi would then allow faster overall decay rates (higher metabolic rates overall) and so more heat output from that metabolism (remember that all respiration generates 'waste' heat). 'Forcing' the system back towards nitrogen limitation by adding more low-N material (like woodchips -- or any 'pure' carbohydrate -- sugar will do!) would force back towards poorer, slower, more fungal-dominated system and less heating.
Some suggested water could play a role. It might -- but be careful about how you make the argument. It might be that bacterial decay is more limited by water availability than fungal decay metabolism, and much the same logic would apply. Properly reasoned, there's a viable hypothesis here, too, but you have to propose that there's a change in limiting resource (You're also missing the rather broad hint about proteins and amino acids. Using water to cool down a pile directly could, of course, work if you kept pouring it on -- but as long as it's 'running hot' with bacterial decay dominant, it'll just heat up again as soon as you stop. Turning the pile or aerating it can have complicated effects, turning may cool briefly, but MOSTLY it tends to allow FASTER decay because it brings more oxygen to the decay organisms -- so actually tends to make piles get hotter and work faster...)
5.
Many
New
England forests have been harvested at intervals of 70-80 years for
over 200 years. The following graph shows general living biomass trends for
such a forest over this time period, with four logging episodes.
You can think of this curve as showing the accumulation of
NPP --
the excess of GPP over respiration and decay, accumulating as biomass
(or as carbon). The
sharply descending parts of the curve show removal of biomass (wood)
in logging. Describe
any other patterns or trends you see over the several cycles of
logging and regrowth. Offer a
hypothesis explaining the dominant patterns in terms of ecosystem
processes; use terms
and concepts from
ecosystem ecology (e.g., you
might need to refer to gross and/or net production, respiration,
limiting resources,...). There's an appearance of
unsustainability here; offer two
possibilities (derived from your hypothesis) for ‘improving’ the
situation; how might
you change things to keep the biomass available for harvest from
becoming less each time?
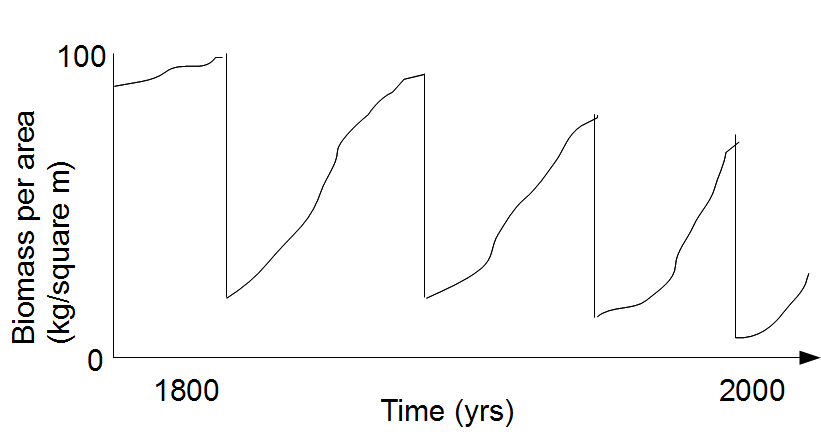
Pattern:
each logging episode (sharp drops in biomass) is followed by
re-accumulation of biomass -- when new growth (NPP) exceeds loss of biomass
through death and decay -- over several decades. This increase,
in at least first cycle, begins to level off, suggesting that, at
ecosystem level, respiration (of all organisms) is 'catching up' with
gross primary production. However, each successive episode of logging
reduces residual biomass to progressively lower levels, and recovering
biomass never reaches levels achieved prior to earlier cuts.
Logging episodes appear to be getting somewhat closer together. It may
be that recovery is somewhat slower at outset in later cycles.
Hypotheses:
Could just be that earlier harvesting with continued removal of large
amounts of biomass (both plausibly driven by economic interests) mean
that regrowth gets a slower start each time (fewer trees left for
seed-source, smaller trees removed), so each interval there's less new
biomass after a given interval -- and economic returns drive continued
intensification of harvest, so residual is less each time -- a
destructive feedback. However, the slower regrowth could also be
a sign of declining fertililty -- decreasing availabilty of some
limiting resource (presumably a mineral nutrient, since CO2 and water
and light availability would not decrease -- if anything, wate rand
light might increase -- following logging). THis could be due to
direct removal (the wood taken contains nutrients that can't be
returned to the soil through decay), or by indirect effects of logging
(like soil erosion).
How to fix it: If first hypothesis is right,
then some combination of simply waiting longer between harvests and
taking less wood (not reducing residual biomass to progressively lower
levels) might be all that it would take. If there is loss of
critical/limiting resources (and there almost has to be some), then the
time between harvests would have to be long enough to allow replacement
of lost nutrients through accumulation of inputs (from weathering of
rock, atmospheric deposition, whatever -- remember the Hubbard Brook story). OR, you could simply add
nutrients actively (fertilize) to replace lost limiting resources
(which is what we'd think of first in a standard ag system).